En primer lugar, los moluscos más evolucionados son los cefalópodos. Pueden medir desde unos pocos centímetros hasta más de 10 m. Los más grandes suelen vivir en zonas más profundas que los más pequeños.
El cuerpo de los cefalópodos presenta una simetría bilateral, en ellos se distingue una cabeza con tentáculos y un pie que utilizan para desplazarse por el método de la retropropulsión. Poseen una concha tabicada la cual puede ser externa como en los ammonites y nautilos o interna como es el caso de los belemnites.
La clase cephalópoda se divide en estas subclases:
-Endoceratoidea
-Actinoceratoidea
-Nautiloidea
-Bactritoidea
-Ammonoidea
-Coleoidea
En este apartado trataré de las subclases Nautiloidea, Ammonoidea y Coleoidea.
Subclase Nautiloidea
Son el grupo más antiguo y duradero de los cefalópodos, ya que aparecieron durante el Cámbrico y hoy en día aún existen en la especie de Nautilus pompilius, la cual vive en los mares del sureste asiático. El estudio del actual Nautilus pompilius nos sirve para poder comprender como eran las partes blandas que no se han fosilizado de los extintos ammonoideos. Los primeros nautiloideos tenían una concha recta similar a la de los belemnites pero externa, pero según ha ido evolucionando la especie, se han curvado las conchas hasta quedar como los actuales. Su rádula, o lengua dentada exclusiva de algunos moluscos, se divide en varias hileras de un gran diente central y seis pequeños a cada lado de este, sumando 13 dientes por fila.
Se alimentaban de crustáceos, tanto grandes como pequeños. Sacaban sus tentáculos en todas las direcciones y según tocaban una presa la traían rápidamente hacia su rádula. Se sabe que engullían los crustáceos con el caparazón, posiblemente porque era una fuente de calcio que usaban para construir su concha. Tenían una pobre visión monocular, y probablemente viesen muy borroso.

Arriba: imágenes de Endoceras sp, un nautiloideo cámbrico de concha ortocónica.
Una cosa extraña en muchos nautiloideos con concha ortocónica era que las cámaras cerca del vértice de la concha, no estaban rellenas de gas, sino de calcita, y según iba creciendo el animal, más cámaras se iban rellenando de calcita, compensándose con las nuevas que este iba desarrollando. Estas cámaras sólidas actuaron como contrapesos para el cuerpo principal del nautiloideo.
Durante el Paleozoico y Mesozoico, otros muchos grupos de nautiloideos aparecieron, pero estos tenían conchas enrolladas en vez de rectas.
Muchas de esas conchas eran muy compactas, y tenían el centro de flotabilidad y de masa juntos. Esto les hacía muy inestables, pero sencillos de manejar, aunque cualquier fuerza movería la concha hacia arriba y abajo.
En contraste, las conchas ortocónicas tenían los centros de masa y flotabilidad separados, serían muy estables y resistentes a los cambios de corrientes, aunque complicados de manejar.
Los nautiloideos de concha enrollada se diversificaron durante el Paleozoico y Mesozoico, mientras que los nautiloideos con concha ortocónica se volvieron menos comunes. Probablemente los nautiloideos de concha ortocónica no pudieron competir con la rápida evolución de los peces óseos y los tiburones, que combinaban velocidad y agilidad, por lo que las últimas especies de nautiloideos de concha ortocónica se extinguieron en el período Triásico. Durante el Mesozoico, los nautiloideos (ya sólo de concha enrollada) se extendieron considerablemente pero siempre menos diversificados que los ammonoideos. Por otra parte, la gran extinción de finales del Cretácico arrasó a los ammonoideos y apenas afectó a los nautiloideos.
Subclase ammonoidea
 -Anarcestida
-Anarcestida
La concha de los ammonites se divide en dos zonas, el fragmocono que es la parte tabicada de la concha donde se almacenaban los gases que controlan la flotación del animal, llenándolo de aire para pesar menos cuando quería emerger y llenándolo de agua para pesar más y descender, igual que hacen los submarinos.


Anarcestina son los primeros ammonites, aparecen en el Devónico temprano y son descendientes directos de los bactrítidos. Tienen líneas de sutura muy simples y desaparecen al final del Devónico.
Clymeniina está representado solamente en el Devónico tardío, y difiere de los demás grupos en la posición del sifúnculo, que es más interior de lo normal.







Fotografía del actual Nautilus pompilius.
Los primeros nautiloideos aparecieron en el Cámbrico temprano, como el Plectronoceras, que tenía una concha cónica ligeramente curvada, similar a la de las lapas. Aunque una concha tan poco hidrodinámica no debería de estar bien adaptada para nadar, la concha del Plectronoceras, tenía un sifúnculo (tubo que conectaba el cuerpo interno y las cámaras del animal con el exterior y echaba agua fuera de las cámaras para rellenarlas de gas), por lo que se supone que era capaz de controlar su flotabilidad, o a lo mejor se movía suspendido a la deriva como las medusas.
En el Ordovícico, los nautiloideos consiguieron, ya de forma segura, controlar su flotabilidad y adquirieron conchas hidrodinámicas, por lo que se volvieron unos mejores nadadores. Las conchas largas y cónicas se convirtieron en ortocónicas, que redujeron el arrastre y les permitieron alcanzar mayores velocidades con menor esfuerzo.
Del Ordovícico en adelante, los nautiloideos tenían una muesca en el borde inferior abriendo la concha (hyponomic sinus), lo cual significó que habían evolucionado a un sistema de propulsión a chorro, aún empleado entre los cefalópodos actuales. Para moverse, absorbían un poco de agua desde el exterior de la concha hasta en interior y la expulsaban apretando fuertemente. El agua salía por un tubo ajustable exclusivo de los nautiloideos (hyponome). En los fósiles actuales, siempre que en un nautiloideo se observa en la parte inferior un pequeño tubo (hyponome), se sabrá que ese nautiloideo se movía por propulsión a chorro y era posterior al período Ordovícico.
Una peculiaridad de la propulsión a chorro, es que el animal se desplaza en la dirección contraria a la que se envía el agua. Como el tubo que expulsaba el agua (hyponome), era flexible, permitía al nautiloideo controlar la dirección a la que quería ir, aunque por otra parte, al curvarlo, se reduciría su eficiencia.
Los nautiloideos actuales, se mueven hacia delante muy lentamente, y su respuesta de escape es un rápido movimiento hacia atrás.

 Al tener una flotabilidad neutral, adquirían un peso inferior, por lo que los nautiloideos con concha ortocónica fueron los primeros "grandes" animales. La mayoría medían entre 30 cm y 2,5 m, mayores que ningún otro invertebrado de esa época. Aunque, se han encontrados fósiles de nautiloideos de nautiloideos con esta concha de más de 5 metros.
Al tener una flotabilidad neutral, adquirían un peso inferior, por lo que los nautiloideos con concha ortocónica fueron los primeros "grandes" animales. La mayoría medían entre 30 cm y 2,5 m, mayores que ningún otro invertebrado de esa época. Aunque, se han encontrados fósiles de nautiloideos de nautiloideos con esta concha de más de 5 metros.
En el Ordovícico, los nautiloideos consiguieron, ya de forma segura, controlar su flotabilidad y adquirieron conchas hidrodinámicas, por lo que se volvieron unos mejores nadadores. Las conchas largas y cónicas se convirtieron en ortocónicas, que redujeron el arrastre y les permitieron alcanzar mayores velocidades con menor esfuerzo.
Del Ordovícico en adelante, los nautiloideos tenían una muesca en el borde inferior abriendo la concha (hyponomic sinus), lo cual significó que habían evolucionado a un sistema de propulsión a chorro, aún empleado entre los cefalópodos actuales. Para moverse, absorbían un poco de agua desde el exterior de la concha hasta en interior y la expulsaban apretando fuertemente. El agua salía por un tubo ajustable exclusivo de los nautiloideos (hyponome). En los fósiles actuales, siempre que en un nautiloideo se observa en la parte inferior un pequeño tubo (hyponome), se sabrá que ese nautiloideo se movía por propulsión a chorro y era posterior al período Ordovícico.
Una peculiaridad de la propulsión a chorro, es que el animal se desplaza en la dirección contraria a la que se envía el agua. Como el tubo que expulsaba el agua (hyponome), era flexible, permitía al nautiloideo controlar la dirección a la que quería ir, aunque por otra parte, al curvarlo, se reduciría su eficiencia.
Los nautiloideos actuales, se mueven hacia delante muy lentamente, y su respuesta de escape es un rápido movimiento hacia atrás.
Al tener una flotabilidad neutral, adquirían un peso inferior, por lo que los nautiloideos con concha ortocónica fueron los primeros "grandes" animales. La mayoría medían entre 30 cm y 2,5 m, mayores que ningún otro invertebrado de esa época. Aunque, se han encontrados fósiles de nautiloideos de nautiloideos con esta concha de más de 5 metros.Arriba: imágenes de Endoceras sp, un nautiloideo cámbrico de concha ortocónica.
Una cosa extraña en muchos nautiloideos con concha ortocónica era que las cámaras cerca del vértice de la concha, no estaban rellenas de gas, sino de calcita, y según iba creciendo el animal, más cámaras se iban rellenando de calcita, compensándose con las nuevas que este iba desarrollando. Estas cámaras sólidas actuaron como contrapesos para el cuerpo principal del nautiloideo.
Durante el Paleozoico y Mesozoico, otros muchos grupos de nautiloideos aparecieron, pero estos tenían conchas enrolladas en vez de rectas.
Muchas de esas conchas eran muy compactas, y tenían el centro de flotabilidad y de masa juntos. Esto les hacía muy inestables, pero sencillos de manejar, aunque cualquier fuerza movería la concha hacia arriba y abajo.
En contraste, las conchas ortocónicas tenían los centros de masa y flotabilidad separados, serían muy estables y resistentes a los cambios de corrientes, aunque complicados de manejar.
Los nautiloideos de concha enrollada se diversificaron durante el Paleozoico y Mesozoico, mientras que los nautiloideos con concha ortocónica se volvieron menos comunes. Probablemente los nautiloideos de concha ortocónica no pudieron competir con la rápida evolución de los peces óseos y los tiburones, que combinaban velocidad y agilidad, por lo que las últimas especies de nautiloideos de concha ortocónica se extinguieron en el período Triásico. Durante el Mesozoico, los nautiloideos (ya sólo de concha enrollada) se extendieron considerablemente pero siempre menos diversificados que los ammonoideos. Por otra parte, la gran extinción de finales del Cretácico arrasó a los ammonoideos y apenas afectó a los nautiloideos.
Subclase ammonoidea
Los ammonoideos (comúnmente denominados ammonites), son los cefalópodos fósiles más abundantes y conocidos, tal es su abundancia que son usados como fósiles guía, es decir, para la datación de rocas. Se conocen más de 1500 géneros de ammonites.
Se dividen en los siguientes órdenes:
-Clymeniida
-Goniatitida
-Prolecanitida
-Ceratítida
-Phyllocerátida
-Lytocerátida
-Ammonítida
-Ancylocerátida
Los ammonites aparecieron en el período devónico y se extinguieron misteriosamente junto con los dinosaurios hace 65 millones de años en el cretácico superior.
La teoría más aceptada de su desaparición es la de el impacto de un meteorito en Chicxulub, península de Yucatán, México, el cual produjo un cambio climático que a su vez produjo la desaparición del placton marino y dejo sin alimento a los ammonites y estos se extinguieron.
Los ammonites eran de diferentes tamaños, podían medir desde unos centímetros hasta más de un metro.
Lo que mejor se conoce de los ammonites era su concha, que estaba compuesta por una capa fina de aragonito, que es una forma de carbonato cálcico.
Así debería de ser un ammonite.
Los ammonites aparecieron en el período devónico y se extinguieron misteriosamente junto con los dinosaurios hace 65 millones de años en el cretácico superior.
La teoría más aceptada de su desaparición es la de el impacto de un meteorito en Chicxulub, península de Yucatán, México, el cual produjo un cambio climático que a su vez produjo la desaparición del placton marino y dejo sin alimento a los ammonites y estos se extinguieron.
Los ammonites eran de diferentes tamaños, podían medir desde unos centímetros hasta más de un metro.
Lo que mejor se conoce de los ammonites era su concha, que estaba compuesta por una capa fina de aragonito, que es una forma de carbonato cálcico.
Esta concha era planiespiralada y con una ornamentación marcada que podía ser de cuatro tipos básicos:
-Rayas: era simplemente una parte de la concha coloreada que podía estar que haciendo formas. Como las rayas rojas del Nautilus actual. Rara vez se encuentran fosilizados junto a la concha pequeños pigmentos de color.
-Espinas: eran pinchos huecos, conectados en un principio con una cámara interior de la concha, como si fuesen una prolongación de esta. Si una espina se rompía el animal no perdía gas por esa zona ni se reducía ligeramente su flotabilidad, pues las espinas, a pesar de estar inicialmente conectadas con las cámaras, se aislaban de estas al desarrollarse, cerrando de nuevo la cámara. Según crecía la concha del ammonite, las espinas se movían hacia atrás en dirección hacia la parte más vieja de la concha y alejándose de la apertura por la que asomaban los tentáculos. Las espinas hacían parecer al ammonite más grande ante sus depredadores e impedían que la concha pudiese ser mordida cómodamente, haciéndola ardua de tragar.
-Quilla: una especie de cadena que sobresalía y recorría a la concha por su parte intermedia exterior, dando al animal una falsa y pequeña mayor altura.
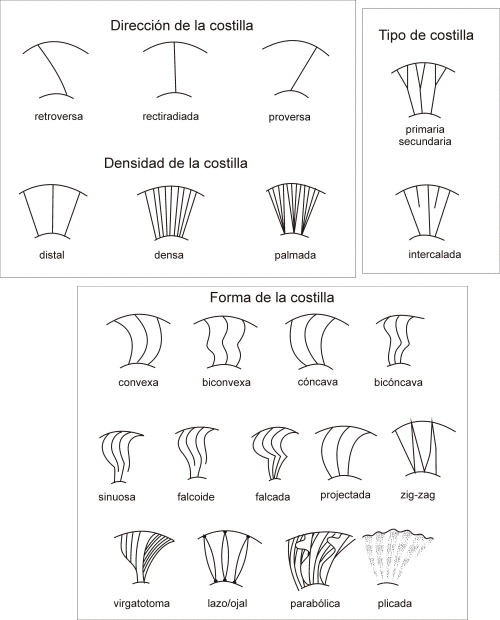
-Costillas: la ornamentación más común de los ammonites, constituída por aros que recorrían la concha a lo ancho y exteriormente. Las habían de muchos tipos según su densidad, dirección o forma:
Aunque estas cuatro no son las únicas decoraciones presentes en las conchas de los ammonites, hay otros cuatro tipos aunque menos importantes:
-Tubérculos: pequeños bultos en la parte más externa de la concha del animal.
-Clavos: los mismos bultos que en los tubérculos pero más alargados.
-Nodos: son como costillas pero que no dan la vuelta a un enrollamiento del ammonite, sólo están por la parte delantera.
-Bulas: son costillas erosionadas y, por cada "costilla" en la parte interior del enrollamiento del ammonite, hay varias en la parte exterior.
La otra parte de la concha de los ammonites es la cámara interior, que es el lugar donde se alojaban las partes blandas del animal. En el interior del fragmocono se encuentran los septos, que tabican esta parte de la concha y tienen su concavidad dirigida hacia la cámara interna. Los septos poseen un orificio, llamado gollete sifonal o sifúnculo, el cual no era poroso, orientado hacia la región posterior que permite la comunicación con el resto de septos a través del sifón.
La intersección de los septos con la pared externa del fragmocono dibuja en ellas la llamada sutura. Existen varios tipos de suturas, los cuales son:
-Ortoceratítica: simple con una pequeña inflexión.
-Agoniatítica: con inflexiones más marcadas y redondeadas.
-Goniatítica: las inflexiones son puntiagudas.
-Ceratítica: inflexiones con forma de herradura.
-Ammonítica: las inflexiones están muy recortadas y con múltiples subdivisiones.
La ventaja directa de la complicación de la sutura está relacionada con el aumento de resistencia de la concha. Cuanto más contacto hay entre el tabique y la pared del fragmocono, más resistente será esta. Esta adaptación les sirvió para poder nadar a mayores velocidades más que para profundizar en el mar, ya que se cree que los ammonites vivían generalmente en zonas costeras y poco profundas.
La línea de sutura también daba a la concha una mayor resistencia frente a fuerzas aplicadas en un punto, distribuyendo estas equitativamente por la concha. Por ejemplo, esto les sería de gran ayuda contra mordiscos, ya que son básicamente fuerzas concentradas en un punto. Sin la sutura compleja, la concha podría implosionar, lo cual sería fatal para el animal, debido a la presión del agua o a fuerzas concentradas en un punto, como ya hemos mencionado. La mayoría de los cefalópodos actuales con concha tabicada no pueden descender a demasiada profundidad debido la presión del agua, en especial las sepias, y teniendo como única excepción a Spirula, que resiste el esfuerzo gracias a su tamaño de 2-3 cm y sus gruesos tejidos que los rodean la concha interna.
Para estudiar el ambiente que habitaban las diferentes especies de ammonites hay que estudiar las posiciones del centro de gravedad y flotación de la concha, ya que estos determinan la forma de desplazamiento. La longitud de la cámara de habitación determina la posición del centro de gravedad, ya que para que el animal flotase, siempre el centro de gravedad o de masa debe estar abajo y vertical respecto al centro de flotación. Esto nos dice que la posición normal de muchos ammonites sería con la apertura algo inclinada hacia arriba, pero al ponerlo en práctica, esta inclinación se vería compensada por el agua expulsada por el sistema de propulsión a chorro, que haría descender la abertura de la concha girando está hacia atrás y variando la inclinación a una posición más normal y que les permitía ver el fondo marino en busca de presas. Esta posición sería con la abertura algo más baja respecto a la posición central. Todo esto, lo veremos ahora con más amplitud:
-Estabilidad estática: está en relación directa con la distancia entre los centros de flotación y gravedad.
-Estabilidad dinámica: depende de la forma de la cámara de habitación, básicamente de la distancia entre la abertura de la concha (punto de empuje del individuo) y el centro de flotación. Gracias a esta características, visibles en las conchas fósiles, se puede determinar si se trataba de buenos o malos nadadores lo que los restringe a un medio marino determinado. Los malos nadadores vivirían principalmente en los fondos marinos y cerca de la costa mientras que los buenos nadadores vivirían en mar abierto.
Se cree que, al igual que la mayoría de los cefalópodos, los ammonites se desplazaban por propulsión a chorro. Cogían agua por el hyponome, lo metían por el sifúnculo y a su vez a otra cavidad, donde lo comprimían y al quedarse el agua sin espacio, salía rápidamente de vuelta por el sifúnculo, y al ser expulsada de nuevo por el hyponome, proporcionaba un gran impulso al ammonite, lo cual le servía repetitivamente para poder moverse. Esta teoría, aunque es la más aceptada, no está realmente confirmada, ya que hay algunos ammonites que debido a sus decoraciones externas (quillas, espinas...), presentan mucha resistencia al mecanismo de propulsión a chorro, restándole a este eficiencia. Este movimiento debería de ser muy rápido, ya que en la cavidad en la cual se comprime el agua no cabría suficiente líquido como para que pudiesen hacer una compresión lenta y larga. Aunque últimamente, se está cuestionando la teoría de la propulsión a chorro aún más, ya que a diferencia de los nautiloideos, los ammonoideos no han sido nunca encontrados con hyponome.
Para desplazarse también es importante ser hidrodinámico y presentar la menor resistencia al movimiento posible. Los ammonites debían vencer la fuerza de la fricción, que les impedía nadar muy rápido si tenían la concha con demasiados salientes, la propia adhesión que presenta el agua sobre este, aunque en seres de tamaño considerable, como en los ammonites, era insignificante, en animales de pequeñas dimensiones esta es fortísima. Por ejemplo cuando un mosquito cae a una piscina y no puede salir de esta porque el propio agua parece que la retiene. La otra fuerza a la que deben de hacer frente al nadar es la succión, esto es el agua que ocupa el espacio que vas dejando atrás mientras te vas moviendo. Pues sólo para mantener el movimiento debían vencer estas tres fuerzas. Si quería mantener una velocidad fija o acelerar, el animal debía ser hidrodinámico.
Como muchas conchas de ammonites no eran demasiado hidrodinámicas, los paleontólogos se están planteando si eran unos buenos nadadores en general, como hemos dicho antes, lo cual les restringía a un medio. Por lo que, a grandes rasgos, los ammonites con conchas en forma de disco y lisas serían mejores nadadores que los que tuviesen conchas globosas y ásperas, o con excesivas decoraciones externas, como costillas o quillas.
Un factor también muy importante que un ammonite debía desarrollar es la defensa. Frente a los predadores este podía quedarse quieto y soportar sus ataques o evitar ser visto y esconderse.
Para soportar los ataques de estos, lo básico que debía de hacer un ammonite era tener una concha gruesa, pero esto aumentaba su masa y reducía su movilidad, por lo que muchos ammonites lo que hicieron fue estrechar zonas de la concha y ensanchar las zonas donde más ataques recibían en forma de una gruesa barrera llamada variz, de la cuales tenían varias a lo largo de la concha.
También algunos ammonites desarrollaron espinas, no ya decorativas, sino defensivas. De esta forma resultaban ser un bocado desagradable para sus captores y dañarían a los que necesitasen meterse toda la concha en la boca para partirla. Además, otros ammonites poseían espinas solo para aparentar un mayor tamaño.
La otra forma de defenderse es escondiéndose, algunos trataban de confundir su contorno gracias a sus colores o a la luz, que es conocido como contrasombreado. Por ejemplo los Nautilus actuales, aunque no son ammonites, tienen unas rayas rojas sobre su concha blanca; estas se concentran arriba de esta y desaparecen casi por completo abajo, de modo que visto desde arriba se confunde con el fondo marino y visto desde abajo no se distingue bien con la luz de la superficie.
También usaban el método del camuflaje, desarrollando estampados o colores en sus conchas. Los más comunes eran en forma de zig-zag y espirales, que se confunden fácilmente, y los colores, aunque se desconocen debido a que estos no se conservan en los fósiles, se cree que los más adecuados serían tonos azules, verdosos e incluso parduscos, siempre esto condicionado al medio en el que vivían, pues no es lo mismo habitar en mar abierto que en la zona costera o en fondos someros.
Anarcestina son los primeros ammonites, aparecen en el Devónico temprano y son descendientes directos de los bactrítidos. Tienen líneas de sutura muy simples y desaparecen al final del Devónico.
Clymeniina está representado solamente en el Devónico tardío, y difiere de los demás grupos en la posición del sifúnculo, que es más interior de lo normal.
Los goniatites (goniatitina) tenían la concha muy involuta con un ombligo muy pequeño. Las líneas de sutura tienen una forma características en zig-zag. Son propios del Paleozoico.
Prolecanitina aparece en el Devónico y se extinge en el Triásico, con conchas achatadas y suturas complejas.
Prolecanitina aparece en el Devónico y se extinge en el Triásico, con conchas achatadas y suturas complejas.
Los ceratites (ceratitina) aparecen en el Triásico y tienen su forma moderadamente evoluta con sutura de líneas redondeadas con dientes. Descienden de Prolecanitina.
Los phylloceras (phylloceratina), lytoceras (lytoceratina) y ammonites (ammonitina), presentan líneas de sutura muy compleja, con sillas y lóbulos. Los phylloceras tienen concha muy involuta con ombligo pequeño, los lytoceras son evolutos y su concha presenta un desarrollo tangencial, mientras que el resto, los ammonites, tienen una gran variedad de formas tanto en el desarrollo de la concha como en sus líneas de sutura. Llaman la atención por sus formas variadas los denominados heteromorfos(ancyloceratina), que presentan formas desenrolladas a los que la espiral no se toca; ejemplos pueden ser los hamites, crioceratites, baculites...
Los dos grandes tipos de conchas tanto para los ammonoideos como para los nautiloideos se definen por el número de veces que se enrolla el espiral alrededor del centro u ombligo, pudiendo ser involutas o evolutas.
-Involutas: la concha sólo se enrolla alrededor del centro una vez. Son las conchas más típicas dentro de los nautiloideos modernos. Dentro de las involutas se distinguen otros dos grupos:
-Los oxiconos: han sido comprimidos lateralmente por lo que son muy finos de perfil.
-Los cadiconos: han sido comprimidos verticalmente y son muy gruesos de perfil.
-Evolutas: la concha se enrolla más de una vez sobre sí misma. Son las más comunes en casi todos los ammonoideos. Dentro de las evolutas se distinguen dos grupos:
-Los serpenticonos: sus enrollamientos de tocan por su parte externa.
-Los cyrtoconos: sus enrollamientos no llegan a tocarse por su parte externa.
Arriba izquierda: concha involuta.
Arriba derecha: concha evoluta.
La alimentación en los ammonites se basaba en presas no excesivamente rápidas y sencillas de cazar debido a que no eran buenos nadadores, por lo que descartamos a los peces. Además, no poseían ganchos ni puntas en los tentáculos como otros cefalópodos. Raras veces se han encontrado ammonites que tenían en su estómago su última comida. Su alimentación por lo tanto sería:
-Organismos bentónicos, como erizos de mar, ostrácodos, crinoideos y algunos foraminíferos.
-Organismos de su propia especie (canibalismo) y otros equinodermos más móviles.
Los ammonites, al igual que la mayoría de la fauna móvil marina, hacían migraciones diarias verticales, lo que significa que al amacencer descendían a aguas más profundas y al anochecer ascendían a aguas más someras.
Visto lo cual, lo ammonites con concha turricónica (de caracola), al ascender por la noche gracias a su concha giraban mientras subían en espiral, cubriendo todos los ángulos ante predadores y extendiendo sus tentáculos para atrapar presas. Todo esto debido a que, como hemos visto antes, al tener que estar necesariamente el centro de masa verticalmente debajo del centro de flotabilidad, ascendían con la concha recta, o sea con la punta de esta mirando hacia la superficie y por lo que la propulsión a chorro la recibirían directamente hacia el fondo marino, haciendo girar a esta sobre sí misma y desplazandole hacia la superficie.
Respecto a su reproducción, se cree que desovaban numerosos huevos de reducido tamaño, y estos se alimentaban del plancton hasta que estaban lo suficientemente formados para la vida adulta. A diferencia de los coleoideos, se cree que los ammonites tenían una vida larga (se estima entre 4 y 36 años) en la que se podrían reproducir varias veces; ya que por ejemplo en los calamares, después de reproducirse muere el macho y al poco la hembra.
Las crías de los ammonites se denominan ammonitelas; dentro del huevo, van formando una protoconcha que luego desarrollaran. El manto de estas segrega el aragonito. Según van llegando a la edad adulta, crecen más despacio, aunque nunca dejan de hacerlo. Es por esto por lo que calculando el número de cámaras del ammonite se puede aproximadamente estimar su edad al morir.
Otro hecho importante a destacar es el dimorfismo sexual. Se cree que la hembra era considerablemente más grande que el macho y se denomina macroconcha.
Por consiguiente, el macho se denomina microconcha, y tiene unas alargaciones en la apertura hacia fuera denominadas orejeras. Estas podían servir para el apareamiento, al igual que ocurre en los pavos reales, y el macho más vistoso se quedaría con la hembra.
El aptychus
El aptychus es una extraña estructura compuesta por dos valvas presente en los ammonites. Ha tenido numerosas dataciones desde su descubrimiento, llegando a considerarse un ser vivo independiente debido a que su gran parecido a un bivalvo y a que está compuesto por calcita, mientras que la concha de los ammonites es de aragonito. También hubo confusiones causadas por el punto en el que son encontradas en los yacimientos, ya que corrientemente no se encuentran cercanos a la concha de los ammonites, porque era una de sus partes peor unidas.
Actualmente existen dos teorías, ambas aceptadas, sobre el aptychus.
Una dice que son las partes de la mandíbula de los ammonites, por su parecido con el de algunos cefalópodos actuales. Si así fuese, serían unas mandíbulas muy desproporcionadas respecto a los cefalópodos de hoy en día.
La otra cuenta que simplemente era una especie de trampilla que cerraban cuando se sentían amenazados mientras soltaban la tinta para despistar a sus perseguidores.
En las dos teorías, la posición del aptychus respecto a la concha es igual.
Antes de que existiese el aptychus, existía el anaptychus, formado por únicamente una valva.
A grandes rasgos, el anaptychus lo poseían los ammonoideos anteriores al Jurásico y los del Jurásico o posteriores tenían el aptychus, formado ya por dos valvas.
Un importante rasgo a destacar es que no todos los ammonites tenían aptychus.
Arriba izquierda: posición del aptychus respecto a la concha del ammonite.
Arriba derecha: aptychus fósil separado de la concha del ammonite.
Subclase coleoidea
Está formada por los actuales órdenes dibranchiata (con dos branquias), que son: Sepiida, Teuthida y Octopódida. Tienen una rádula igual a la de los ammonites, un gran diente central y cuatro a cada lado (nueve dientes por hilera).
También está formada por los órdenes extinguidos de Aulacoceridea, Phragmoteuthida y Belemnítida. En este apartado sólo trataré del órden Belemnítida.
Los belemnoideos se caracterizan por tener una concha interna muy parecida a la de los actuales calamares. Son fáciles de encontrar en los estratos del Jurásico y del Cretácico, a veces formando grandes acumulaciones de estos conocidas como campos de batalla belemníticos. Los belemnites más pequeños no superan el centímetro, pero los más grandes, como el Megateuthis, su rostro llega a medir hasta un metro y medio.
El rostro es la parte fosilizada que se puede encontrar hoy en día, y es lo que formaba su esqueleto interno. Su cuerpo se divide en un rostro sólido y en un rostro hueco.
Este último forma el alveolo, el cual contiene el fragmocono.
Arriba: campo de batalla belemnítico.
Abajo: partes de un belemnite.














{kind=link}